
一文详解—双荧光素酶报告基因技术
发布时间:
2025-03-21 09:26
来源:
双荧光素酶报告基因是以萤火虫荧光素酶(Firefly luciferase)为报告基因,以海肾荧光素酶(Renilla luciferase)为内参基因,通过同时检测两种荧光素酶,实现对目标基因表达或分子互作的定量分析。
检测原理
将研究的调控元件(如启动子、miRNA靶序列等)插入带有萤火虫荧光素酶(Firefly luciferase)的表达载体,构建成报告基因质粒,使这段序列调控萤火虫荧光素酶的转录表达。然后将报告基因质粒转染细胞,并加入底物荧光素(luciferin),由于萤火虫荧光素酶可催化荧光素发出荧光(最强波长在560 nm左右),通过检测得到的荧光值高低,即可判断不同处理组下荧光素酶的活性,进而定量分析目标基因表达或分子互作。
同时将海肾荧光素酶基因作为内参使用,将两个报告基因构建到同一个质粒上,分别用不同的启动子启动其表达,消除细胞转染效率和细胞转录活性对实验结果产生的影响。
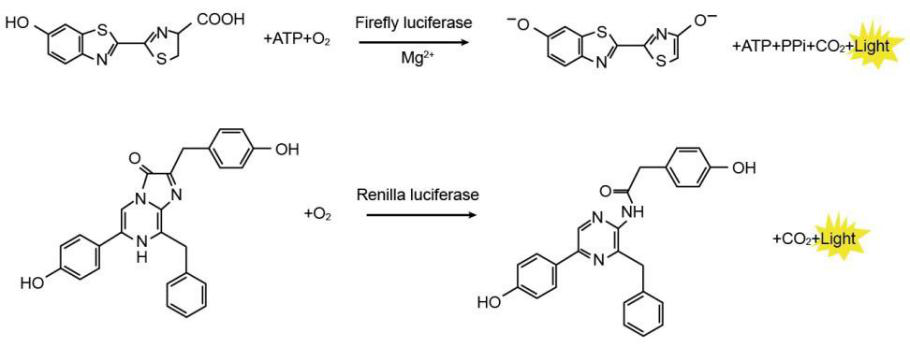
图1:双荧光素酶报告基因原理
主要应用
1. 验证miRNA与靶基因的作用
miRNA可以通过基因剪切或翻译抑制等方式来调控靶基因,精准控制其表达水平。通过将待测靶基因序列(3’UTR)插入LUC的报告基因载体,当miRNA能够和靶基因作用时,miRNA会通过和所插入的序列结合从而抑制荧光素酶的翻译使荧光值降低。
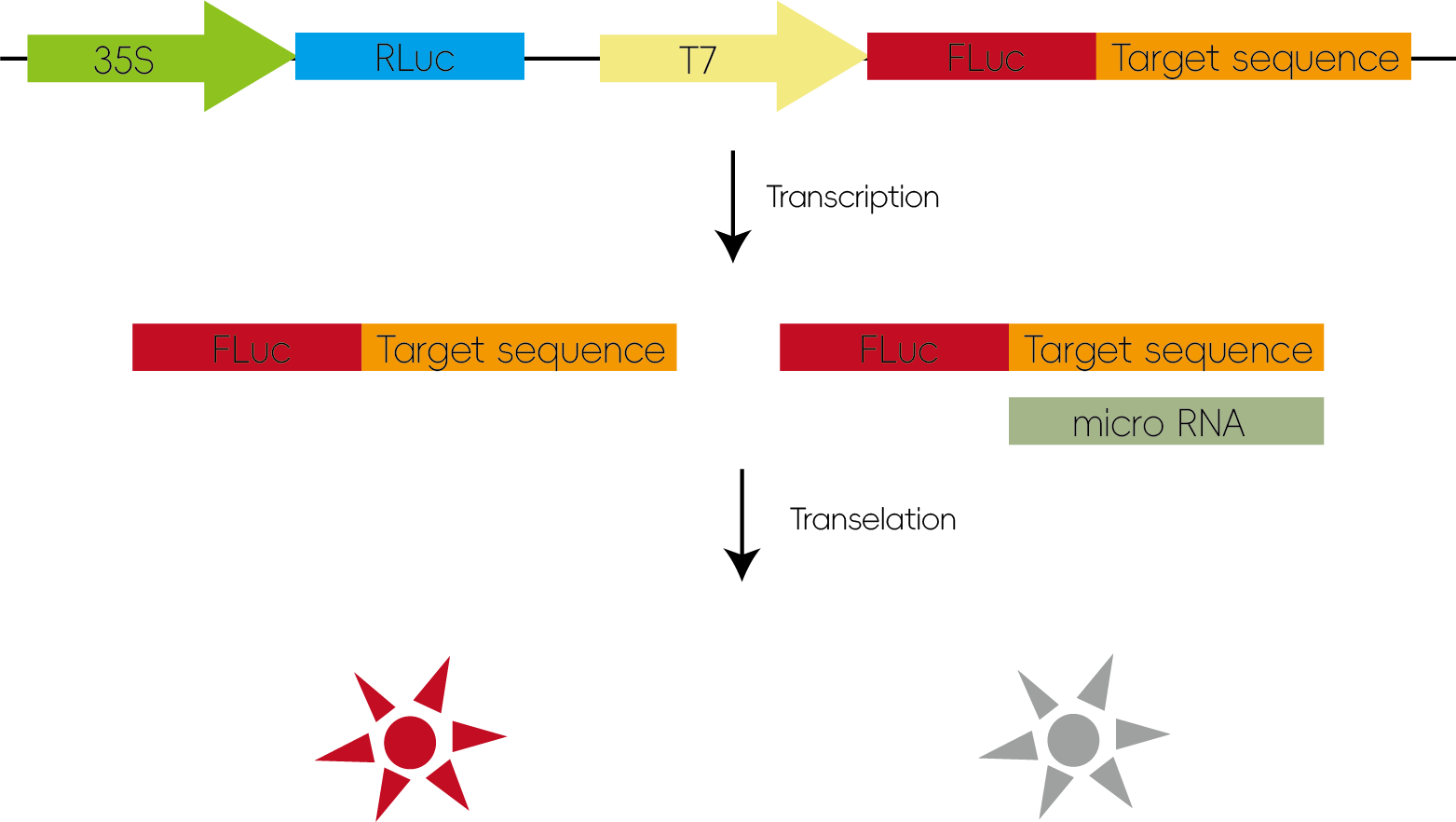
图2:验证miRNA与靶基因的作用
2. 研究miRNA和lncRNA/circRNA的靶向互作
许多lncRNA具有与mRNA相似的结构,miRNA可通过与作用于mRNA类似的机制对lncRNA起负调控作用。将候选的lncRNA序列插入报告基因载体上,当miRNA能够和Inc/circRNA作用时,miRNA会通过和所插入的序列结合从而抑制荧光素酶的翻译使荧光值降低。
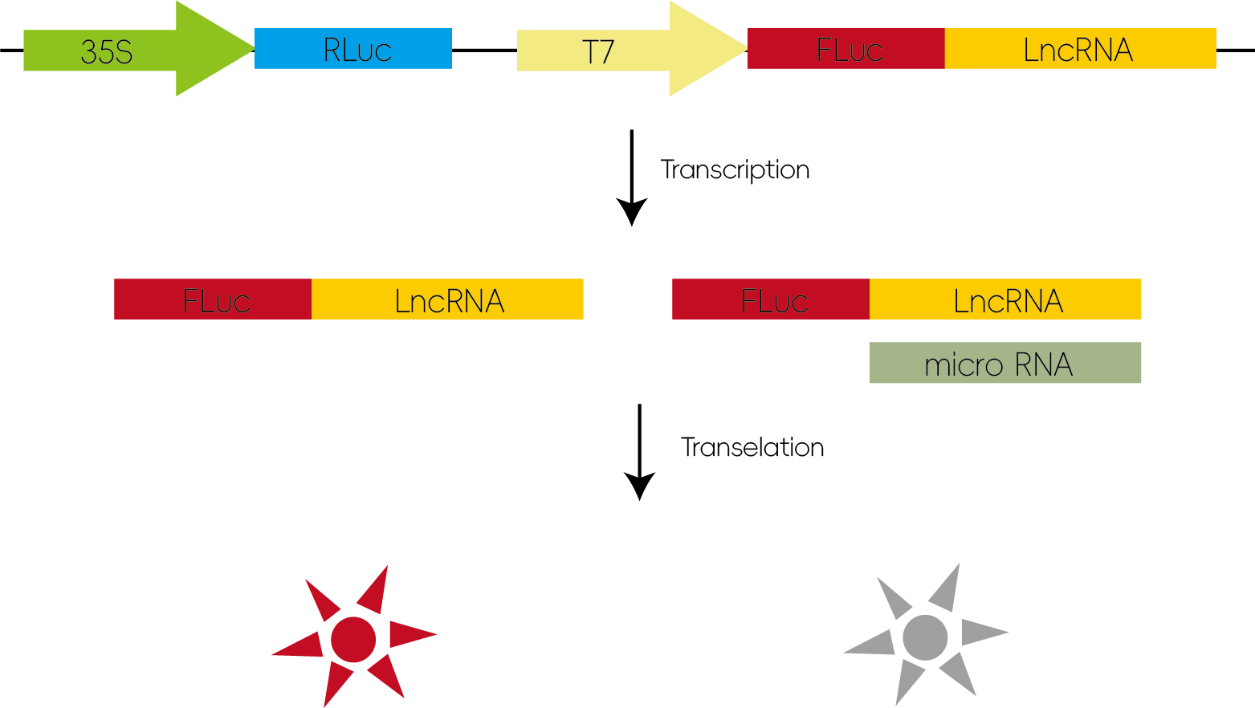
图3:研究miRNA和lncRNA/circRNA的靶向互作
3. 研究转录因子对下游基因的作用
转录因子与其靶启动子中的特异序列(顺式作用元件)结合,从而对基因的表达进行调控。通过将启动子区域序列插入LUC的报告基因载体,同时共转入转录因子,当转录因子能与对应启动子结合,则可以转录翻译产生荧光素酶提升荧光值。
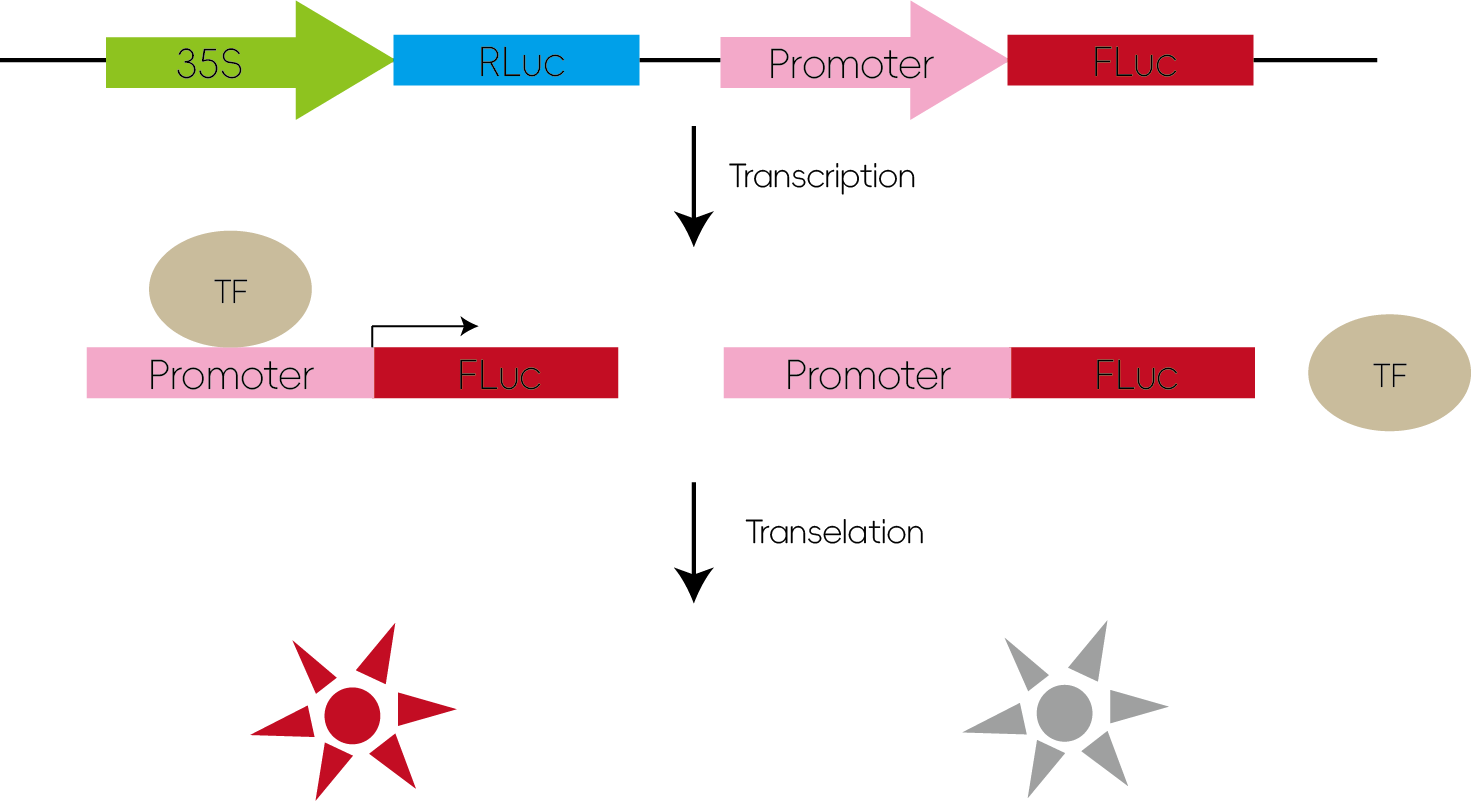
图4:研究转录因子对下游基因的作用
4. 启动子结构分析
将启动子区域序列进行分段截短,或对特定位点进行突变,再分别构建入荧光素酶报告载体,检测其启动子活性。
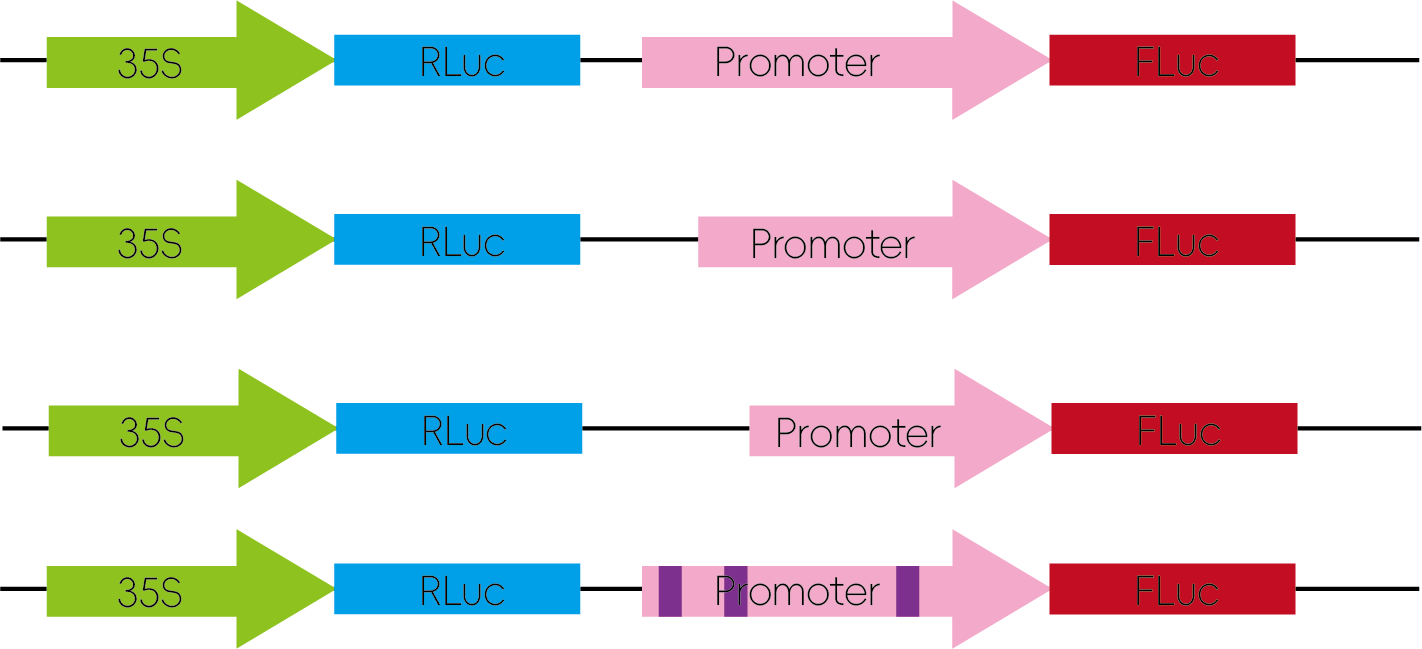
图5:启动子结构分析
5. 启动子SNP分析
一些基因的启动子区域存在单核苷酸多态性,可运用萤光素酶报告系统分析其相对活性。

图6:启动子SNP分析
6. 验证启动子活性
将待测启动子构建至荧光素酶基因的上游,检测启动子的活性。

图7:验证启动子活性
7. 验证转录因子活性
将待测转录因子与GAL4结合域构建在同一载体上,与含有GAL-TATA转录调控原件并带有荧光虫荧光素酶(Firefly luciferase)的表达载体共转,使这段序列调控luciferase的转录表达。通过检测荧光值的高低可以判断转录因子是否具有转录激活或者转录抑制作用。
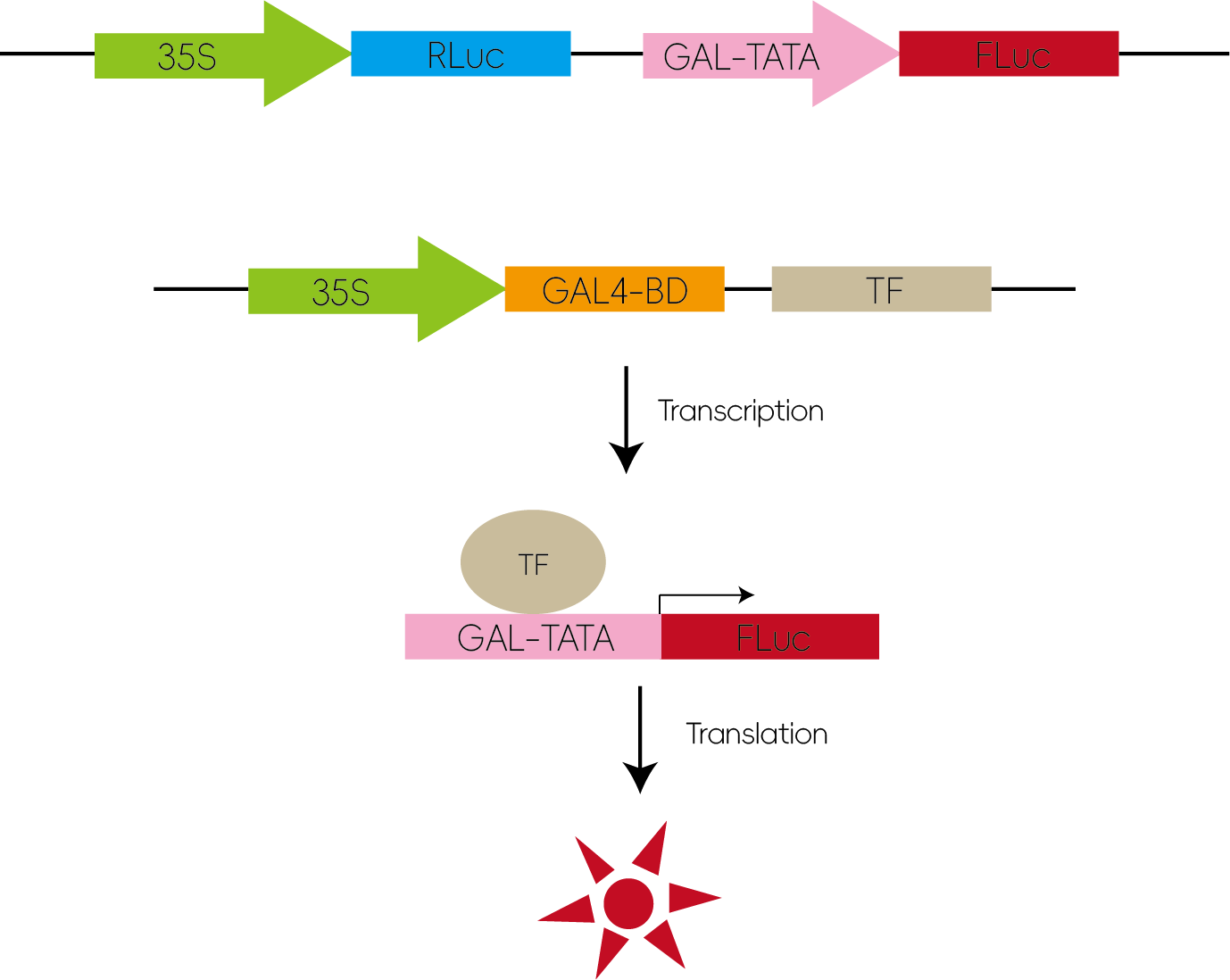
图8:验证转录因子活性
产品优势
样品要求

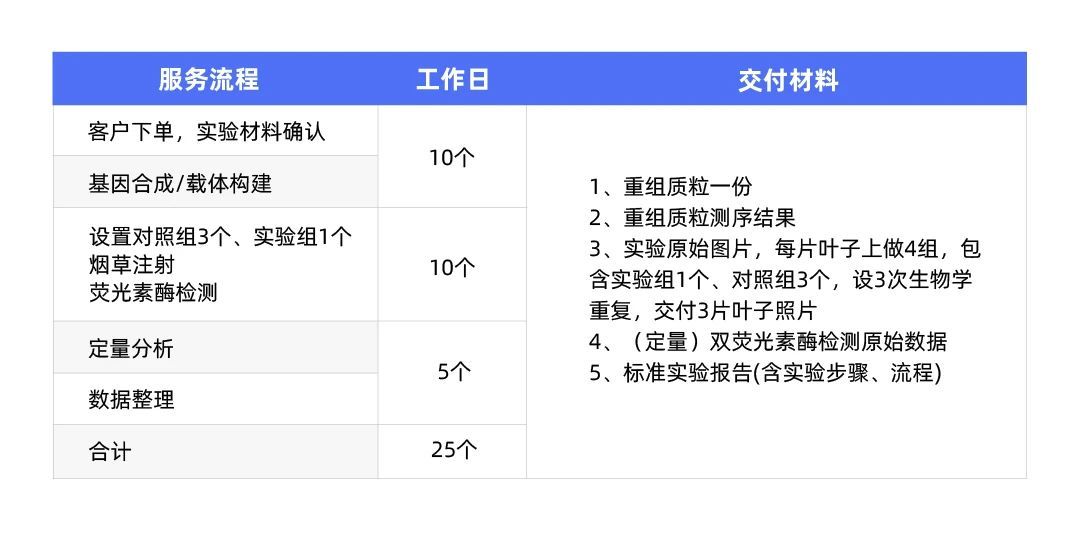
服务内容

案例展示
2021 年 10 月 7发表于New Phytologist ( IF=8.3 )上一篇题为ZmMPK5 phosphorylates ZmNAC49 to enhance oxidative stress tolerance in maize的研究型论文种,作者将ZmSODs(ZmSOD1、ZmSOD2、ZmSOD3、ZmSOD4)的启动子序列克隆至载体p1381-LUC中。效应载体使用玉米泛素启动子(Ubi)驱动ZmNAC49表达(Ubi:ZmNAC49),以空载体作为对照。结果发现ZmNAC49激活了ZmSOD3启动子的LUC表达,表明ZmSOD3是ZmNAC49 的靶基因。

图9:ZmSODs与ZmNAC49互作研究
2020年发表于Molecular Plant(IF= 17.1)上一篇题为“Novel miR167a-OsARF6-OsAUX3 Module Regulates Grain Length and Weight in Rice”的研究型论文,作者通过生物信息学分析发现OsARF6的编码区存在miR167a互补序列。于是作者将OsARF6的野生型3'UTR(含miR167a结合位点)克隆至荧光素酶报告载体,构建pGREEN-OsARF6:LUC。
同时将同义突变破坏miR167a结合位点,构建突变型报告载体pGREEN-mOsARF6:LUC。然后作者将miR167a前体与野生型/突变型报告载体共转染水稻原生质体。设置四组比较:空载体 ± miR167a前体(基线对照);野生型OsARF6:LUC ± miR167a前体;突变型mOsARF6:LUC ± miR167a前体。结果发现野生型组miR167a前体使荧光素酶活性显著降低而突变型组miR167a前体无法抑制荧光素酶活性,于是作者得出结论OsARF6与miR167a存在互作,且该互作为特异性的。
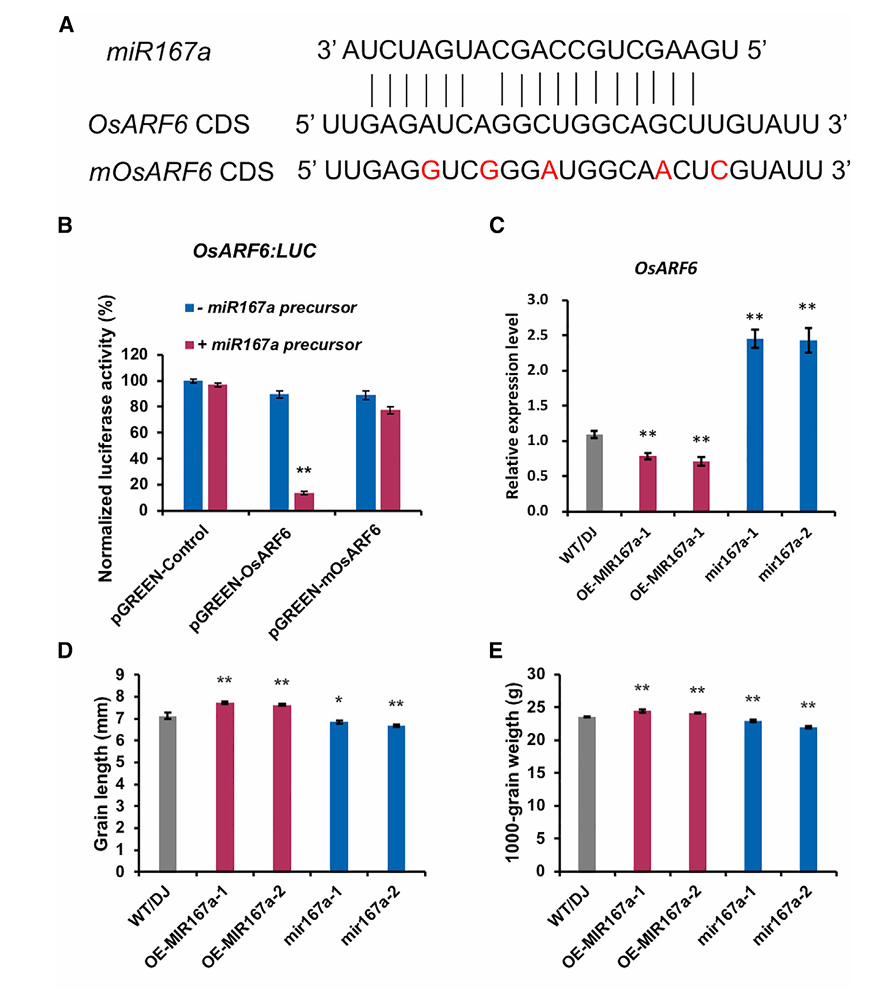
图10:OsARF6与miR167a的互作研究
常见问题

相关新闻
双荧光素酶报告基因是以萤火虫荧光素酶(Firefly luciferase)为报告基因,以海肾荧光素酶(Renilla luciferase)为内参基因,通过同时检测两种荧光素酶,实现对目标基因表达或分子互作的定量分析。

赋能科研,驱动创新
联系我们
-
电话:400-863-8520
咨询邮箱:ruiyuanyx@bestofbest.top
投诉建议:info@bestofbest.top
地址:江苏省南京市栖霞区仙林街道齐民西路6号南京仙林智谷3C栋10楼
子公司:南京维百瑞检测技术有限公司


南京瑞源生物
官方客服